
1.
Реально до підігрівників ЕК буде прикладатися напруга на 1,5 У нижче розрахункового за рахунок падіння цієї напруги на відкритому тиристорі V6. У моїй схемі до підігрівника реально прикладається напруга 22,5 У.
2.
За рахунок природного підвищення величини опору нагрівального елемента при його розігріві токовище через підігрівник зменшується. Реальне токовище через один підігрівник ЕК становить 0,7 А проти 0,96 А з розрахунку.
За рахунок цих двох причин реальна потужність підігрівника становить 16 Вт проти 23 Вт із розрахунку.
Ці особливості роботи схеми треба обовязково враховувати при розрахунку реальної потужності підігрівника.
Якщо потужність виготовленого підігрівника виявиться недостатньої, то її можна збільшити або за рахунок збільшення питающегонапряжения, що знімається із трансформатора, або за рахунок зменшення опору (довжини) нагрівального елемента. У нашім варіанті для збільшення реальної потужності нагрівача до 20 Вт треба або збільшити живильна напруга, що знімається із вторинної обмотки, до 30 У (якщо така можливість є), або відрізати близько 5 Ом опору, обмірюваного на «холодній» дроті.
І ще одне зауваження. Підігрівник забезпечує задану потужність при номінальному значенні живлячої напруги мережі в 220 У. Однак, з ряду причин, особливо в сільській місцевості, напруга в мережі може підтримуватися нижче номіналу. В окремих місцевостях напруга мережі буває навіть менш 200 У. Тому, якщо виявиться, що не вистачає потужності підігрівника, пошук причини треба починати з виміру номіналу живлячої напруги мережі.
У схемі як випрямляч використовується мостова схема на діодах V1 -V4 типи КД 203Б. Однак можуть використовуватися й інші силові діоди на токовище 5-10 А. На транзисторі VT1 зібрана схема стабілізації для харчування мікросхеми. Виконавчим елементом схеми терморегулювання є тиристор V6 типу КУ 202 з будь-яким буквеним індексом від Г до Н. Діод V5 виконує функцію розвязки напруги, що прикладається до тиристора й підігрівників. Діод V5 може бути будь-якого типу на токовище не менш 0,1 А. Лампа Н1 і светодиоды V7, V8 використовуються для контролю роботи схеми терморегулювання.
> Силовий блок для обігріву приміщення
Обігрів приміщення виробляється від мережі 220 У без трансформатора. Випрямний міст блоку зібраний на силових діодах V1 -V4. Це можуть бути будь-які діоди на токовище 10 А с зворотною напругою не менш 300 У. Виконавчим елементом схеми є тиристор V7 типу КУ 202Н(М).
Як нагрівальний елемент можна використовувати будь-які побутові электрорадиаторы, электроконвекторы, електроплитки закритого типу потужністю 500—1 000 Вт. Якщо температура поверхні нагрівача буде більше 80—90 °С, то цю поверхню треба накрити товстим металевим листом. Зазначеної потужності нагрівача досить для підтримки необхідної для зимівлі температури 5 ± ГС у приміщенні обсягом до 10—12 м з теплими (деревяними, з пінобетону, керамзиту й т.д.) стінами й добре утепленою стелею. У приміщенні більшого обсягу потужність нагрівача з даним силовим блоком можна збільшити, але не більш ніж до 2 кВт. При цьому силові діоди V1 ^4і особливо тиристор V7 повинні перебувати на вертикально розташованих металевих радіаторах площею 40—50 див кожний. Светодиоды V5, V6 використовуються для контролю роботи схеми.
Если буде ухвален рішення здійснювати автоматичний обігрів тільки в приміщенні й не займатися внут-риульевым підігрівом, то в цьому випадку треба буде виготовити тільки силовий блок для обігріву приміщення й блок терморегулювання з окремим блоком живлення для нього.
Блок живлення в цьому випадку виготовляється за аналогією із блоком живлення для підігріву вуликів. Але трансформатор Т1 може мати потужність не більше 5-10 Вт, вторинна напруга 26-30 У. Діоди V1 -V4 будь-які на токовище не більше 0,1 А. Силовий вузол - тиристор V6, светодиоды V7, V8 і відповідні опори виключаються.
Також виключається діод V5. Вихідна напруга цього блока живлення виставляється підбором типу стабілітрона або ланцюжка стабілітронів у ланцюзі бази транзистора VT1 і може становити від 27 до 30 У. Може використовуватися й будь-який інший блок живлення зі стабілізатором на відповідну напругу й токовище не менш 0,1 А.
Увага! При складанні схеми терморегулювання для приміщення треба мати на увазі, що всі елементи силового блоку будуть перебувати під небезпечним для життя потенціалом 220 У. Під цим же потенціалом будуть перебувати й елементи блоку терморегулювання. Із цієї причини всі ці елементи повинні бути надійно ізольовані від корпусів цих блоків і недоступні для випадкового
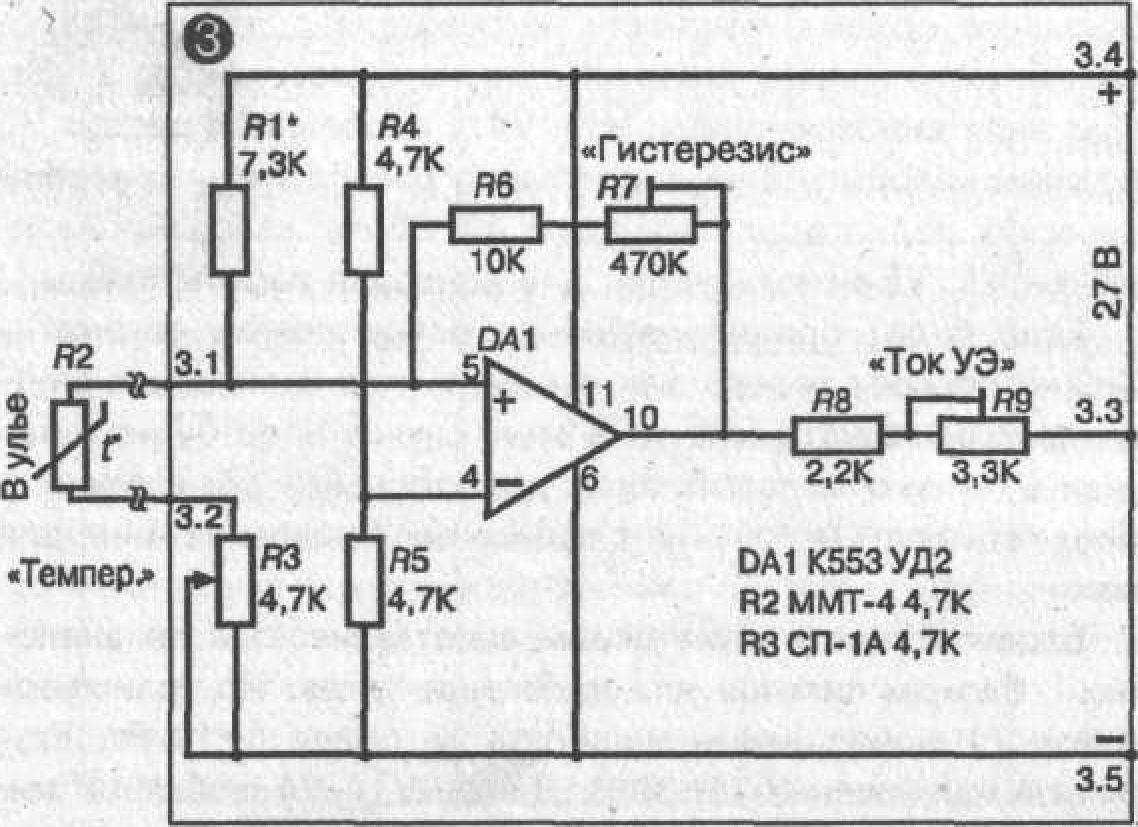
Рис. 3.37. Принципиальная схема блока терморегулировадотику при роботі. _ _ _
ния
► Блок терморегулювання.
Блок терморегулювання зібраний на мікросхемі ДО553УД2. Термочувствительным елементом (термодатчиком) є термоопір R2 4,7 кому типу ММТ-4, що міститься в один з вуликів над поверхнею підігрівника під низ рамок.
При настроюванні схеми потенціометром R7 «Гістерезис» виставляється ширина температурної петлі гістерезису в 1, 5-2 °З. Робиться це так: обертається ручка потенціометра R3 «Температура» в одну сторону до спрацьовування схеми. На шкалі «Температура» відзначається крапка 1 спрацьовування (включення) схеми. Потім потенціометр R3 обертається в протилежну сторону й відзначається крапка 2 зворотні спрацьовування (вимикання) схеми. Відстань між крапками 1 і 2 у °С и є ширина температурного гістерезису. Регулюванням R7 домагаються, щоб різниця цих температур була не більше 1,5—2 °С.
Потенціометром R1 «токовище УЭ» виставляється токовище керуючого електрода, при якому відбувається надійне спрацьовування (відкриття тиристора). При цьому треба перевірити, чи буде при виставленому токовищі УЭ відбуватися закриття тиристора. У моїй схемі для тиристора КУ 202Л виставлений /
= 8 мА.
Підбором R1 виставляється значення обраного температурного діапазону автоматичного регулювання на шкалу «Температура», що перебуває на передній панелі блоку. При зазначених на схемі значеннях опорів діапазон автоматичного регулювання блоку лежить у межах від 0 до 50 "С. Градуювання шкали «Температура» виробляється за допомогою надійного термометра. Термодатчик R2 і термометр поміщають у місце з фиксируемой температурою на кілька мінут. Необхідно мати на увазі, що теплова инерционность термоопору в повітряному середовищі становить не менш 120 секунд. Не меншу инерционность має й термометр. Тому для надійного фіксування температури термоопір і термометр треба тримати в середовищі з фиксируемой температурою не менш 8—10 мінут. Після цього обертанням ручки потенціометра R3 домагаються вимикання электроподогревателя ЕК і на шкалі «Температура» робиться відповідна оцінка. Температура, що у цей час показує термометр, буде відповідати зробленої на шкалі оцінці. Звертаю увагу на те, що шкала «Температура» повинна градуюватися не на включення підігрівника, а на його вимикання. Справа в тому, що для однієї й тої ж температури, за рахунок теплової петлі гістерезису, положення ручки на шкалі «Температура» при включенні й вимиканні підігрівника буде відрізнятися на 1,5—2 °С.
Зазначене вище регулювання блоку означає, що при працюючій схемі терморегулювання у вулику буде підтримуватися температура не вище тої, котра буде виставлена на шкалі.
У якості фиксируемых температур можна користуватися кімнатною температурою, температурою над поверхнею добре прогрітого электроподогревателя, температурою усередині холодильника, температурою льоду, що тане (0 С) і ін. У ході експлуатації апаратури градуювання шкали «Температура» уточнюється.
Для можливості вибору можна порекомендувати також оригінальну схему регулятора температур, що наведена в № 9 журналу «Бджільництво» за 2005 рік на стор. 44.
Закінчуючи розгляд питання, хочеться дати пораду тим, хто збирається виготовити комплект апаратури для терморегулювання: перш ніж почати виготовлення, обовязково прочитайте ту частину додатка 3, у якій говориться про способи підключення навантажень.
І останнє. Звертаю увагу на те, що представлена схема терморегулювання може з успіхом використовуватися в побуті й для інших цілей: регулювання температури в інкубаторах, акваріумах і т.д.
Дивлячись на дерево, що скинуло листя,, многим з нас здається, що життя в ньому зупинилася до.
наступної весни. Дехто нітрохи не сумнівається в цьому, однак не всі так просто й однозначно в тих процесах, які відбуваються в такому, здавалося б, відомому нам рослині, як дерево...
Учені багатьох країн, що вивчають життєдіяльність (фізіологію) деревних рослин, поки не прийшли до єдиної думки щодо того, як саме деревний організм реагує на зниження температури навколишнього середовища, які процеси при цьому й у якій послідовності відбуваються в капілярах і живих клітинах деревини, чому й при яких умовах відбувається повне припинення життєдіяльності (вимерзання) рослин.
При глибокому проробленні цього питання виникає безліч приватних проблем. От тільки один приклад - вода. Уже, здається, що може бути простіше!.
У багатьох з нас ще зі шкільного років міцно встановився стереотип про те, що вода замерзає при температурі 0 "С. Однак виявляється, що сама звичайна вода, що перебуває в скляній трубочці з мікрокапіляром в 0,1 мм, замерзає при —20 °С (Чудинов Б.С., 1984). У той же час у деревному капілярі такого ж діаметра вона замерзає при більше високій температурі. Прийнятного пояснення цьому факту поки ніхто не дав. Більше того, є инфррмация про те, що за певних умов звичайна вода може залишатися в рідкому стані (так звана переохолоджена вода) при температурах аж до -70 °С (Веретенників А:В., 1987).
Існує також і ряд інших проблем, по яких поки немає однозначного розуміння.
По всіх цих причинах пропонована модель функціонування живого дерева листяної породи в період часу осінь - зима - весна не може претендувати на безперечність. Однак, з обліком всієї наявної інформації із цього питання на сьогоднішній день, ця модель представляється найбільш імовірної.
Процес підготовки дерева до зими починається задовго до її настання. Уважається, що механізм попередньої підготовки дерева до зими включається з початком скорочення світлового дня, коли в дереві починають припинятися процеси росту. Потім наприкінці літа дерево вступає в стан так званого внутрішнього спокою, при якому в деревному організмі різко знижується інтенсивність обміну речовин.
При подальшому стійкому зниженні температури повітря нижче +10 °С, найчастіше це відбувається в серпні, дерево переходить у стан глибокого (органічного) спокою, що для більшості деревних рослин середньої смуги триває до листопада — грудня (Сергєєв Л.И., 1964). По іншому джерелу - до листопада - січня (Лісова енциклопедія, 1986).
Восени з настанням холодів обмінні процеси в дереві продовжують уповільнюватися. Зменшуються у звязку із цим і можливості провідної системи дерева па доставці вологи й живильних речовин до галузей (особливо тонким) і листам. Однак транспірація (випар) вологи в атмосферу не припиняється, і для того щоб зменшити транспірацію й привести її хоч у якусь відповідність із можливостями по доставці вологи, дерево скидає листя. Таким чином, дерево захищає себе від зимового иссушения.
У процесі характерних для осені багаторазових перепадів температур відбувається так зване загартовування дерева. У ході цього періоду усередині живих клітин дерева починають накопичуватися захисні речовини у вигляді розчинів Цукрів і деяких інших речовин. Ці захисні речовини створюють у дереві такі умови, при яких сильно знижується ймовірність утворення льоду усередині живих клітин. Але навіть у найбільш морозостійких дерев вся наявна в тканинах вода не може залишатися незамерзаючої. Якась її частина при тривалому впливі низьких негативних температур все-таки буде замерзати. Саме головне, щоб цей процес відбувався не в протопласті (оболонці) живої клітини, а в міжклітинному просторі, тоді протопласт буде піддаватися меншої небезпеки одержати механічні ушкодження.
У лабораторних умовах при поступовому східчастому зниженні температури вдалося домогтися надзвичайно високої морозовитривалості дерева. Так, в одному з досвідів И.И. Туманова гілка яблуні, поміщена в переносну морозильну камеру з температурою —153 °С, навесні цвіла й навіть плодоносила (по А.А. Веретенникову, 1987).
При подальшому зниженні зовнішніх температур відтік води в міжклітинні простори збільшується; у цей момент часу там буде перебувати рівно стільки води, скільки її зможе кристалізуватися при наявній у цей момент температурі.
Процес загартовування дерева й нагромадження води в межклетниках відбувається поступово й, залежно від характеру наступаючої зими, може займати тривалий період часу. У ході цього періоду сокодвижение в дереві ще відбувається, хоча й в уповільненому темпі.
З переходом зовнішніх температур в область стійких негативних значень корнеобитаемый шар ґрунту поступово промерзає, що приводить до зменшення вологи, що поглинається коріннями.
При подальшому охолодженні деревини наявна в ній вода після невеликого переохолодження починає замерзати спочатку в найбільш великих капілярах. Але властивості води такі, що процес її кристалізації супроводжується виділенням теплоти. Так, при замерзанні одного грама води виділяється 335 Дж (80 кал) тепла. Із цієї причини температура деревини, у капілярах якої починає замерзати вода, підвищується до 0 °С и стабілізується на цьому рівні. Пояснюється дане явище тим, що в лід за одиницю часу може перетворитися рівно стільки води, скільки необхідно для того, щоб теплота, що виділяється при цьому, компенсувала теплоту, що приділяється за цей же час, при охолодженні деревини зовнішнім повітрям. Температура в окремих крапках усередині деревини почне опускатися нижче 0 °С лише після того, як вся вільна вода біля них змерзне. •.
Вода в капілярах дерева (так звана вільна вода) починає замерзати тільки при охолодженні деревини до —2...5 °З (Чудинов Б.С., 1984). Інше джерело затверджує, що в стовбурах деревних порід утворення льоду відбувається при -2...4 "З, а при сухій атмосфері - при -8...11 "З (Самыгин Г.А., 1974).
Однак з початком замерзання в стовбурі кристалізується тільки частина води, а частина цієї вільної (не звязаної в клітинах) води в міжклітинних просторах залишається в рідкому стані й сокодвижение, хоча й дуже уповільнено, триває до ще більш низьких температур. Здатність води (пасоки) зберігатися в переохолодженому рідкому стані в тканинах деревини, наявність у них розчинів органічних і мінеральних речовин, що сприяють збереженню рідкого стану води (на зразок антифризу), своєчасна «загартування» дерева всі перепадами, що збільшуються, температур охороняють дерево від загибелі навіть при тріскучих морозах.
Що ж стосується звязаної в клітинах вологи, то вона починає замерзати при її охолодженні до — 5...—16 °С (Самыгин Г.А., 1974), до -10...15 °З (Чудинов Б.С., 1984). Беручи до уваги значну инерционность температурного поля в деревних стовбурах, особливо великого діаметра, це відбувається, видимо, при зниженні температури в атмосфері до —15...—20 °С (Чудинов Б.С., 1984).
Кількість кристаллизуемой у межклетниках води поступово збільшується, і у випадку настання тривалого періоду низьких негативних температур у грудні - січні дерево переходить у стан змушеного (екзогенного) спокою.
Цей стан характеризується наявністю в міжклітинних просторах заболоні певної кількості замерзлої води, пъчти повною відсутністю сокодвижения в стовбурі й галузях, продовженням кутикулярной (покривної) транспірації й, як наслідок, постійним дефіцитом вологи в живих тканинах дерева. Подих живих клітин, що випробовують дефіцит вологи, триває постійно, хоча й з різко, що зменшилася інтенсивністю. Дерево знизило всі свої жизнеобеспечивающие функції до мінімально можливого рівня, що, однак, достатній для поновлення цих функцій з настанням тимчасового або постійного потепління. Підсушені живі клітини готові в будь-який момент прийняти відсутню воду, що перебуває в міжклітинних просторах у замерзлому й переохолодженому стані, як тільки вона нагріється й перейде в рідкий агрегатний стан.
Паралельно з описаними вище процесами в цей же час у дереві відбувається й інший процес. Як показують дослідження (Крамер П., Козловский Т., 1983), з початком переходу дерева в стан внутрішнього спокою наприкінці літа починає змінюватися концентрація газів у стовбурі дерева: концентрація кисню починає підвищуватися, а вуглекислого газу — зменшуватися. До січня — лютому кисень уже може займати більше 15% загального обсягу стовбура (сосна) і до 30% у тополі, у той час як вуглекислий газ і в сосни й у тополі займає не більше 5% обсягу стовбура. Ці зміни концентрацій газів викликаються змінами обмінних процесів, які у свою чергу визначаються зниженням середньодобових температур. При цьому чітко простежується зворотна залежність між значеннями зовнішніх температур і концентраціями ПР0
і пряма залежність між температурами й концентраціями З
. Наявність цього процесу означає, що обсяг стовбура, наприклад, тополі в зимовий час зайнятий у такому співвідношенні: близько 30% деревиною, близько 30% — вільною й звязаною водою в різних агрегатних станах і більш ніж на 40% — газом (ПРО
+ З
) (Крамер П., Козловский Т., 1983).
Газ у деревині заповнює ті порожнини й порожнечі, які не зайняті водою. Якби газ перебував у капілярах разом з водою й розривав безперервні водні потоки, то це зробило б неможливим пересування водних потоків, тому що обовязковою умовою для пересування рідини в капілярі є безперервність і нерозривність водного потоку.
Наявність у стовбурі зимового дерева великої кількості порожнеч, що становлять до 40% загального обсягу стовбура й зайнятих газом, надає деревині гарні теплоизолирующие властивості.
Що ж стосується можливості поповнення запасів води дрібними галузями в холодну пору року (а саме вони випробовують у цей час найбільший дефіцит вологи), те И.И. Туманів (1955) установив наступне. Зрізані пагони, що зберігалися взимку на відкритому місці, містять менше води, чим пагони, що перебували в тих же умовах на рослинах. Пояснюється це тим, що навіть у холодні місяці відбувається поповнення вологи в галузях за рахунок припливу води з інших частин дерева, де є запаси, вологи. Таке пересування води може бути й при слабких морозах, якщо в деревному організмі залишається ще достатня кількість води в рідкій фазі. Поповнення водного дефіциту можливо навіть при значних морозах. Воно спостерігається при сонячній погоді, коли надземні частини можуть помітно прогріватися в порівнянні з навколишнім повітрям. Таким шляхом тонкі гілочки, що мають більшу поверхню випару, періодично поповнюють витрачену воду за рахунок її запасів у більше масивних частинах рослин. Якщо тривалий час через сильні морози цього відбуватися не буде, то галузі будуть ушкоджуватися й надалі відмирати.
Основний, причиною ушкодження деревини при замерзанні рослин уважається зневоднювання цитоплазми клітини, викликане утворенням льоду в межклетниках. Усе більше й більше відтягаючи воду із цитоплазми, лід збільшується в обсязі й починає чисто механічно ушкоджувати поверхневі шари збезводненої цитоплазми.
Ю.З. Кулагин (1969) також указує на те, що під час зимових відлиг і слабких (менш 5—7 °С) морозів у пагони надходить вода з товстих кістякових галузей і стовбура, що ліквідує небезпечний водний дефіцит, викликаний зимовим иссушением.
Однак при настанні тривалого періоду низьких негативних температур (—15...—20 °С) дерево переходить у стан змушеного спокою, при якому сокодвижение в дереві буде значною мірою уповільнено, а в окремі періоди дуже низьких температур і зовсім припинятися.
Провесною з початком потепління при прогріві стовбура до позитивних температур відновляється інтенсивне сокодвижение, у першу чергу за рахунок запасів вільної вологи, що зберігається зимою в міжклітинних просторах. У міру прогріву ґрунту й корнеобитаемого шаруючи відновляється надходження вологи від корінь, включається механізм активного поглинання води деревним організмом. Рушійною силою активного поглинання є так званий кореневий тиск. Коренева система при цьому перетворюється у своєрідний насос, що жене вологу по стовбурі. Механізм активного поглинання діє тільки ранньої навесні. Надалі дерево переходить на пасивне поглинання вологи за рахунок кутикулярной (з поверхні дерева), а потім - і листовий транспира-ции, що тепер буде основною рушійною силою, що викликає переміщення вологи в провідній системі дерева.
Додаток 2.